- Индийский океан
- Индийский океан
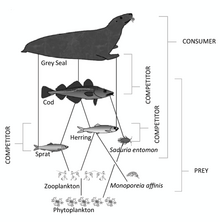
- Краткая промыслово-экологическая характеристика Индийского океана
- Цепи питания индийского океана
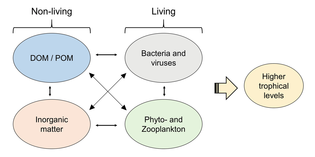
- Морская пищевая сеть — Marine food web
- СОДЕРЖАНИЕ
- Пищевые цепи и трофические уровни
- По таксону
- Первичные производители
- Основные потребители
- Потребители высшего порядка
- Микроорганизмы
- Грибы
- По среде обитания
- Пелагические сети
- На поверхности океана
- На дне океана
- Прибрежные сети
- Полярные сети
- Виды фундамента и краеугольного камня
- Загадочные взаимодействия
- Сложность и стабильность
Индийский океан
Индийский океан
Краткая промыслово-экологическая характеристика Индийского океана
В отличие от Атлантического и Тихого, Индийский океан почти целиком расположен в Южном полушарии. Только самые северные участки океана, составляющие лишь пятую часть его акватории, находятся севернее экватора в тропической и экваториальной зонах. На юге Индийский океан ограничен Антарктидой, а его самая южная часть входит в выделяемый некоторыми океанологами самостоятельный Южный океан.
Акватория Индийского океана (76,2 млн. км2 ) несколько меньше акватории Атлантического. Средняя глубина его — 3711 м, максимальная (7209 м) расположена в глубоководном Зондском желобе. Шельф занимает 6,1% всей акватории, что меньше, чем в Атлантическом, но больше, чем в Тихом океане. Почти все моря и заливы Индийского океана находятся в его северной и восточной частях (Аравийское море с Аденским, Оманским и Персидским заливами; Бенгальский залив, Андаманское и Красное моря). У побережья Австралии расположены: Арафурское и Тиморское моря, а также Большой Австралийский залив.
Южнее Африки и Австралии Индийский океан свободно сообщается с Тихим и Атлантическим. Наиболее широкий шельф в Индийском океане занимает северо-восточную часть Аравийского моря, а также располагается вдоль побережья западного Индостана, северного побережья острова Шри-Ланка и на севере Бенгальского залива. Средняя величина первичной продуктивности Индийского океана составляет 170-220 граммов С/м2 в год (почти не отличается от таковой в Мировом океане).
Особенностью процессов биопродуцирования в Индийском океане является ярко выраженная сезонность в развитии апвеллингов в экваториальной и тропической его зонах (в отличие от высокоширотных апвеллингов в Атлантическом и Тихом океанах). Поэтому пищевые цепи здесь обычно короткие и малоэффективные, так как около 30% первичной продукции не включается в повторный цикл и рассеивается. С другой стороны, Индийский океан, в отличие от Атлантического и Тихого, лишен самой биопродуктивной (северной высокоширотной) зоны, что снижает его общую биологическую продуктивность. Указанные специфические особенности Индийского океана приводят к изобилию здесь низших звеньев трофической цепи, например, мелких мезопелагических рыб, причем эти звенья не в полной мере используются консументами высших порядков. Общий запас мелких мезопелагических рыб в Индийском океане оценивается в 350 млн. т. Если в Индийском океане биомасса этой группы составляет 4,57 т/км2 , то в Атлантическом — 1,75, а в Тихом — 2,25.
Однако биоресурсы Индийского океана слабее (в основном по причинам экономико-географического характера) освоены промыслом, чем биоресурсы Тихого или Атлантического. Из 88 млн. т гидробионтов, добытых в 1992 г. в Мировом океане, в Индийском было добыто лишь 7 млн. т (в Атлантическом — при примерно той же акватории — 23,8 млн. т, в Тихом — 51,4 млн. т), то есть лишь около 9%. Основой рыбного промысла в Индийском океане являются скомброидные рыбы (скумбрии, тунцы и др.), которых здесь добывается около 1 млн. т в год (1992 г.), ставридовые (314 тыс. т), сельдевые (сардинелла с годовым выловом около 300 тыс. т), горбылевые (около 300 тыс. т), акулы и скаты (около 170 тыс. т в год).
Промысловая статистика ФАО ООН подразделяет Индийский океан на три региона: западную часть (ЗИО), восточную (ВИО) и Антарктическую (АЧИО). Западная часть Индийского океана включает Аравийское море, Персидский залив, а также восточные шельфы Африки и прилегающие участки открытой части Индийского океана, включая воды Мальдивских, Сейшельских, Коморских, Амирантских и Маскаренских островов, а также Маврикия и Мадагаскара. В Аравийском море наиболее биопродуктивны зоны прибрежных апвеллингов, в частности, прибрежные воды Аравии и западного Индостана. По величине первичной продукции это одни из наиболее биопродуктивных районов Мирового океана. Воды Аравийского моря наиболее благоприятны для пелагических рыб, которые здесь составляют 71% в уловах в среднем. Главным объектом про-67 мысла является индийская сардинелла, которой здесь добывают около 100 тыс. т ежегодно (в основном кустарными орудиями лова).
Персидский залив расположен целиком внутри материковой отмели. Хотя здесь имеется богатая промысловая ихтиофауна (тунцы, мечерылые, акулы, ставридовые, сельдевые, мелкие скумбриевые и др.), добыча рыбы здесь не играет большой роли, так как прибрежные государства (Иран, Ирак, Кувейт, Саудовская Аравия, Бахрейн, Катар, Объединенные Арабские Эмираты) занимаются в основном нефтедобычей. Годовой улов рыбы здесь не превышает 7090 тыс. т. В водах Мальдивских островов главный объект рыболовства — тунцы, из которых 70% составляет пятнистый тунец. 80% тунцов, акул и мечерылых, добываемых в открытых водах Индийского океана, вылавливается Японией и Южной Кореей, некоторую долю добывают российские тунцеловы и мальдивские рыбаки.
В западной части Индийского океана имеются промысловые скопления мезопелагических рыб (светящихся анчоусов рода Diaphus) Восточная часть Индийского океана (ВИО) включает Бенгальский залив, воды Андаманских и Никобарских островов, воды, прилегающие к западному побережью островов Суматра и Ява, шельф северной и западной Австралии, Большой Австралийский залив и прилегающие воды открытой части Индийского океана. В северной части Бенгальского залива имеется широкий шельф. Во всей прибрежной зоне залива есть ряд апвеллингов нестационарного характера. Кроме того, на формирование высокой биологической продуктивности залива оказывает мощное влияние сток одной из крупнейших рек азиатского континента — Ганга, опресняющее влияние которого прослеживается на всей акватории залива (воды Бенгальского залива имеют соленость 26-34 промилли).
Ихтиофауна насчитывает 475 видов рыб. Рыболовство наиболее активно ведется в зимний период, когда усиливаются северо-восточные муссонные ветры. Вблизи берегов и эстуариев рек много рыб из семейств анчоусовых, сельдевых, ставридовых, лутьяновых, ворчуновых и горбылевых. К нерито-океанической экологической группе можно отнести ряд представителей скумбриевых, ставридовых, а к собственно океанической — кархариновых акул, скатов, тунцов и др. Промысловые скопления рыб наиболее часто можно встретить на глубинах до 60 м. Это сардинеллы, ящероголовы, анчоусы, пальцеперы, спаровые (морские караси), горбылевые, индийская скумбрия и др. По оценкам специалистов, только у западного побережья Индии можно добывать без ущерба для запасов 8- 12 млн. т рыб. Уже сейчас доля вылова в Бенгальском заливе в отдельные годы составляет до 80% от всего вылова в Индийском океане. Примерно 50% от всего улова составляют пелагические виды — индийская скумбрия и сардинелла, около 30% — пристипома из семейства ворчуновых (помадасиевых). В целом улов рыбы, по мнению ихтиологов, можно увеличить без ущерба для воспроизводства примерно в 10 раз.
В открытых водах восточной части Индийского океана, особенно в островных зонах, ведется промысел тунцов, акул и мечерылых. Большие запасы рыб имеются в водах индоокеанского шельфа Австралии, их запасы оцениваются в 20 млн. т. Особенно рыбопродуктивны воды мелководья северной Австралии. Здесь большие запасы рыб из семейств ворчуновых, ставридовых, сребробрюшковых, сельдевых и др. Промысловый интерес представляют также: Большой Австралийский залив, воды южной Австралии, где имеются большие запасы сардины, анчоуса, скумбрии, ставриды, полосатого и других тунцов.
Открытые воды юго-восточной части Индийского океана менее биопродуктивны, чем юго-западной, так как они содержат меньше биогенных элементов. Антарктические воды Индийского океана (АЧИО). Ихтиофауна этого района представлена 44 видами рыб, относящихся к 16 семействам. Промысловое значение имеют только нототениевые и белокровные рыбы, а также антарктический криль, которые здесь весьма перспективны для промыслового освоения. В целом же биоресурсы этого района беднее, чем биоресурсы антарктической части Атлантического океана.
В открытых водах Индийского океана имеются весьма перспективные для развития промысла скопления акул (146 видов из 21 семейства), наиболее массовые из которых голубая (или синяя) акула, обыкновенная лисья акула, белоперая серая акула. Добываются они обычно как прилов при ярусном промысле тунцов. Из тунцов — объектов ярусного лова — наибольшее промысловое значение имеют желтоперый, большеглазый, длинноперый, австралийский и длиннохвостый тунцы. Кроме того, считается перспективным развитие кошелькового промысла полосатого, индо-тихоокеанского пятнистого тунцов, ауксид и ярусного промысла южного тунца и мечерылых.
Источник
Цепи питания индийского океана
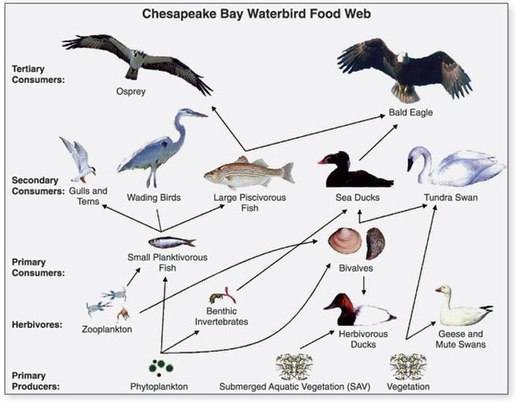
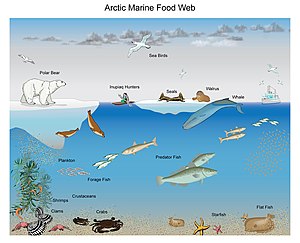
Морская пищевая цепь представляет собой сложную систему, в которой мелкие организмы потребляются более крупными. В нижней части пищевой цепи находятся микроскопические растения, а в верхней части — известные хищники, такие как акулы и морские птицы. В зависимости от их размера и места в пищевой цепи, рыба служит различным целям и помогает сбалансировать экосистему разными способами.
Производители фитопланктона
Основной производитель пищевой цепи называется фитопланктон. Производители создают свою еду. Эти одноклеточные микроскопические растения плавают на поверхности океана, получают энергию от солнца и используют ее для превращения углекислого газа и других питательных веществ в углеводы, которые питают другие виды океана. Фитопланктон, такой как диатомовые водоросли и водоросли, является основой пищевой цепи океана. Они составляют 95 процентов первичных производителей на земле.
Зоопланктон
Зоопланктон — это маленькие, плавающие животные. Они включают личинок рыб, медуз, микроскопических копепод и мелких донных животных. Они дрейфуют через океан и питаются фитопланктоном. Веслоногие ракообразные составляют большинство зоопланктона. Они составляют большую часть массы животных океана и являются наиболее важной связью между основными производителями и многими крупными животными, питающимися планктоном, такими как мелкая сельдь. Почти все рыбы, обитающие в умеренных или полярных водах, едят веслоногих, чтобы выжить в какой-то момент своей жизни.
Маленькие Хищники
Следующий уровень в пищевой цепи состоит из мелких хищников, которые питаются рачками и другим планктоном, который они добывают из воды. Моллюски, мелкие ракообразные, такие как креветки и криль, и мелкие рыбы, такие как сардины и сельдь, питаются большим количеством зоопланктона. Крупные стаи мелкой рыбы могут быстро уменьшить популяции планктона, но только временно.
Лучшие хищники
Крупные хищники, такие как акулы, тунец, кальмар и осьминог, а также морские млекопитающие, такие как тюлени и некоторые киты, образуют вершину пищевой цепи. Птицы и люди также включены в эту группу. Крупные хищники питаются большим разнообразием мелких рыб. Такие виды, как голубая рыба и полосатый окунь, являются не только одними из самых популярных целей для ловли рыбы на людях, но они также потребляются крупной рыбой, такой как рыба-меч и акула, а также скопами и другими морскими птицами, которые захватывают их из воды. Это показывает, как даже рыба на вершине пищевой цепи может стать пищей для других главных хищников. Самые лучшие хищники будут есть все, что доступно, включая друг друга. Омары — одни из самых известных людоедов океана.
Пищевая цепочка начинается снова
Пища, которую тратят эти крупные хищники, дрейфует на дно океана, где его питают омары и другие обитатели дна. Часть пищи также разлагается бактериями и возвращается в почву, где растения могут использовать ее питательные вещества. Трата китов и морских черепах, существ, у которых нет непосредственных хищников, также разрушается бактериями.
Источник
Морская пищевая сеть — Marine food web

| Часть серии обзоров по | |||||||||||
| морская жизнь | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
 | |||||||||||
| |||||||||||
 Портал морской жизни Портал морской жизни | |||||||||||
| морская пищевая цепь (типовая) |
|---|
 солнце солнце |
| ↓ |
| фитопланктон |
| ↓ |
| травоядный зоопланктон |
| ↓ |
| хищный зоопланктон |
| ↓ |
 питатель фильтра питатель фильтра |
| ↓ |
 хищное позвоночное животное хищное позвоночное животное |
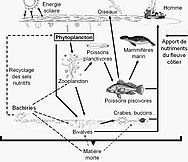
Пищевые сети построены из пищевых цепочек . Все формы жизни в море могут стать пищей для другой формы жизни. В океане пищевая цепочка обычно начинается с энергии Солнца, питающей фитопланктон , и следует таким курсом, как:
фитопланктон → растительноядный зоопланктон → плотоядный зоопланктон → фильтр-питатель → хищное позвоночное
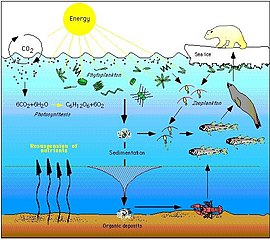
Фитопланктону не нужны другие организмы для пищи, потому что они способны производить себе пищу непосредственно из неорганического углерода, используя солнечный свет в качестве источника энергии. Этот процесс называется фотосинтезом , и в результате фитопланктон превращает природный углерод в протоплазму . По этой причине считается, что фитопланктон является основными продуцентами на нижнем или первом уровне морской пищевой цепи. Поскольку они находятся на первом уровне, считается, что у них трофический уровень 1 (от греческого trophē, означающего пищу). Затем фитопланктон потребляется на следующем трофическом уровне пищевой цепи микроскопическими животными, называемыми зоопланктоном.
Зоопланктон составляет второй трофический уровень пищевой цепи и включает микроскопические одноклеточные организмы, называемые простейшими, а также мелких ракообразных , таких как веслоногие ракообразные и криль , а также личинки рыб, кальмаров, омаров и крабов. Организмы на этом уровне можно рассматривать как первичных потребителей .
В своей очереди, меньшая растительноядная зоопланктона потребляются более крупным хищным зоопланктоном, такие как больший хищный простейших и криль , а кормовая рыбой , которые являются небольшими, школьным обучение , фильтр кормления рыб. Это составляет третий трофический уровень пищевой цепи.
Четвертый трофический уровень — это хищная рыба , морские млекопитающие и морские птицы , потребляющие кормовую рыбу. Примеры: рыба-меч , тюлени и олуши .
Высшие хищники, такие как косатки , которые могут поедать тюленей, и короткоперые акулы мако , которые могут поедать рыбу-меч, составляют пятый трофический уровень. Усатые киты могут напрямую потреблять зоопланктон и криль, что приводит к пищевой цепочке с тремя или четырьмя трофическими уровнями.
На практике трофические уровни обычно не являются простыми целыми числами, потому что один и тот же вид потребителей часто питается более чем на одном трофическом уровне. Например, крупное морское позвоночное животное может есть хищную рыбу меньшего размера, но может также есть кормушки с фильтром; то скат ест ракообразные , но и молоты съедают как ракообразные и скаты. Животные тоже могут есть друг друга; что трески съедает меньше трески, а также раки и раки едят трески личинку. Пищевые привычки молодняка и, как следствие, его трофический уровень могут изменяться по мере взросления.
Ученый- рыболов Дэниел Поли устанавливает значения трофических уровней: один для первичных продуцентов и детрита , два — для травоядных и детритофагов (первичные потребители), три — для вторичных потребителей и так далее. Определение трофического уровня TL для любого вида-потребителя:
Т L я знак равно 1 + ∑ j ( Т L j ⋅ D C я j ) <\ Displaystyle TL_ <я>= 1 + \ сумма _ 
где — фракционный трофический уровень жертвы j , а представляет долю j в рационе i . В случае морских экосистем трофический уровень большинства рыб и других морских потребителей принимает значение от 2,0 до 5,0. Верхнее значение 5,0 необычно даже для крупной рыбы, хотя встречается у высших хищников морских млекопитающих, таких как белые медведи и косатки. Для сравнения: средний трофический уровень человека составляет около 2,21, что примерно соответствует уровню свиньи или анчоуса. Т L j <\ displaystyle TL_ D C я j <\ displaystyle DC_
По таксону
Первичные производители

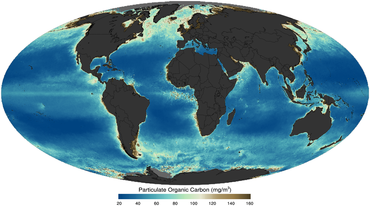
В основе пищевой сети океана лежат одноклеточные водоросли и другие растительные организмы, известные как фитопланктон . Как и растения на суше, фитопланктон использует хлорофилл и другие светособирающие пигменты для фотосинтеза , поглощая атмосферный углекислый газ для производства сахаров в качестве топлива. Хлорофилл в воде изменяет то, как вода отражает и поглощает солнечный свет, что позволяет ученым наносить на карту количество и расположение фитопланктона. Эти измерения дают ученым ценную информацию о состоянии окружающей среды океана и помогают ученым изучать углеродный цикл океана .
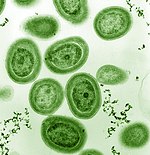
Фитопланктон входит в группу бактерий, называемых цианобактериями . Морские цианобактерии включают мельчайшие из известных фотосинтезирующих организмов. Самый маленький из них, прохлорококк , имеет диаметр всего от 0,5 до 0,8 микрометра. С точки зрения индивидуальной численности, Prochlorococcus , возможно, самый многочисленный вид на Земле: один миллилитр поверхностной морской воды может содержать 100 000 клеток или больше. Во всем мире насчитывается несколько октиллионов (10 27 ) особей. Prochlorococcus повсеместно распространены между 40 ° с.ш. и 40 ° ю.ш. и доминируют в олиготрофных (бедных питательными веществами) регионах океанов. Бактерия составляет около 20% кислорода в атмосфере Земли.
- Фитопланктон составляет основу пищевой цепи океана



В океанах большая часть первичной продукции производится водорослями . Это контрастирует с землей, где основная часть первичной продукции производится сосудистыми растениями . Водоросли варьируются от отдельных плавающих клеток до прикрепленных водорослей , в то время как сосудистые растения представлены в океане такими группами, как морские травы и мангровые заросли . Более крупные продуценты, такие как морские травы и водоросли , в основном ограничены литоральной зоной и мелководьем, где они прикрепляются к нижележащему субстрату и все еще находятся в фотической зоне . Но большая часть первичной продукции водорослей осуществляется фитопланктоном.
Таким образом, в океанской среде первый трофический уровень дна в основном занимает фитопланктон , микроскопические дрейфующие организмы, в основном одноклеточные водоросли , которые плавают в море. Большинство фитопланктона слишком малы, чтобы их можно было увидеть невооруженным глазом . Они могут проявляться в виде (часто зеленого) обесцвечивания воды, когда они присутствуют в достаточно большом количестве. Поскольку они увеличивают свою биомассу в основном за счет фотосинтеза, они живут в освещенном солнцем поверхностном слое ( эвфотической зоне ) моря.
Наиболее важные группы фитопланктона включают диатомовые водоросли и динофлагелляты . Диатомовые водоросли особенно важны в океанах, где, по некоторым оценкам, они составляют до 45% от общей первичной продукции океана. Диатомовые водоросли обычно микроскопические , хотя некоторые виды могут достигать в длину до 2 миллиметров.
Основные потребители
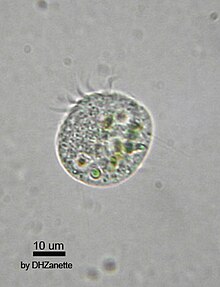
Второй трофический уровень ( первичные потребители ) занимает зоопланктон, питающийся фитопланктоном . Вместе с фитопланктоном они составляют основу пищевой пирамиды, которая поддерживает большинство великих рыболовных угодий мира. Многие зоопланктона крошечные животные нашли с фитопланктона в океанических поверхностных водах , и включают в себя крошечные ракообразные и рыбы личинок и мальков (недавно вылупившиеся рыба). Большинство зоопланктона являются фильтраторами и используют придатки, чтобы напрячь фитопланктон в воде. Некоторый более крупный зоопланктон также питается меньшим зоопланктоном. Некоторые зоопланктоны могут немного прыгать, чтобы избежать хищников, но они не умеют плавать. Как и фитопланктон, они плавают вместе с течениями, приливами и ветрами. Зоопланктон может быстро воспроизводиться, при благоприятных условиях их популяция может увеличиваться до тридцати процентов в день. Многие живут короткой и продуктивной жизнью и быстро достигают зрелости.
В oligotrichs представляют собой группу инфузорий , которые имеют известные пероральные реснички , расположенные как воротник и отворот. Они очень распространены в сообществах морского планктона, обычно в концентрациях около одного на миллилитр. Это самые важные морские травоядные животные, первое звено пищевой цепи.
Другими особо важными группами зоопланктона являются веслоногие рачки и криль . Веслоногие ракообразные — это группа мелких ракообразных, обитающих в океанических и пресноводных средах обитания . Они являются крупнейшим источником белка в море и являются важной добычей кормовой рыбы. Криль — следующий по величине источник белка. Криль — это особенно крупный зоопланктон-хищник, который питается зоопланктоном меньшего размера. Это означает, что они действительно относятся к третьему трофическому уровню, вторичным потребителям, наряду с кормовой рыбой.
- Зоопланктон формирует второй уровень пищевой цепи океана.

![]()
Крошечные креветочные ракообразные
![]()
Молодь планктонных кальмаров

Вместе фитопланктон и зоопланктон составляют большую часть планктона в море. Планктон — это термин, применяемый к любым маленьким дрейфующим организмам, которые плавают в море ( греч. Planktos = странник или бродяга). По определению организмы, классифицируемые как планктон, не могут плавать против океанских течений; они не могут противостоять окружающему току и контролировать свое положение. В условиях океана первые два трофических уровня заняты в основном планктоном . Планктон делится на производителей и потребителей. Производителями являются фитопланктон (греч. Phyton = растение), а потребителями, которые питаются фитопланктоном, является зоопланктон (греч. Zoon = животное).
Медузы медленно плавают, и большинство видов составляют часть планктона. Традиционно медузы рассматривались как трофические тупики, второстепенные игроки в морской пищевой сети, студенистые организмы, строение тела которых в значительной степени основано на воде, которая представляет небольшую питательную ценность или интерес для других организмов, за исключением нескольких специализированных хищников, таких как океаническая солнечная рыба и кожистых морских черепах . Это мнение недавно было оспорено. Медузы и, в более общем смысле, студенистый зоопланктон, который включает сальпы и гребневики , очень разнообразны, хрупки, не имеют твердых частей, их трудно увидеть и контролировать, они подвержены быстрым колебаниям численности и часто живут неудобно вдали от берега или глубоко в океане. Ученым сложно обнаружить и проанализировать медузу в кишечнике хищников, поскольку они превращаются в кашу при поедании и быстро перевариваются. Но медузы цветут в огромных количествах, и было показано, что они являются основными компонентами рациона тунца , рыбы-копья и рыбы-меч, а также различных птиц и беспозвоночных, таких как осьминоги , морские огурцы , крабы и амфиподы . «Несмотря на их низкую плотность энергии, вклад медуз в энергетический баланс хищников может быть намного больше, чем предполагалось, из-за быстрого пищеварения, низких затрат на вылов, доступности и избирательного питания более богатыми энергией компонентами. морские хищники восприимчивы к проглатыванию пластмасс ».
Потребители высшего порядка

В 2010 году исследователи обнаружили, что киты переносят питательные вещества из глубин океана обратно на поверхность с помощью процесса, который они назвали китовым насосом . Киты питаются на более глубоких уровнях океана, где водится криль , но регулярно возвращаются на поверхность, чтобы дышать. Здесь киты испражняются жидкостью, богатой азотом и железом. Вместо того, чтобы тонуть, жидкость остается на поверхности, где ее поглощает фитопланктон . В заливе Мэн китовый насос обеспечивает больше азота, чем реки.
![]()
![]()
Оуши ныряют сверху, чтобы поймать кормовую рыбу

Микроорганизмы

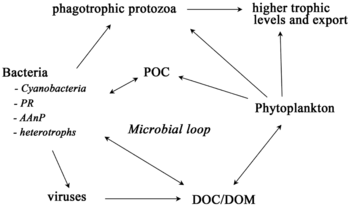
В последние годы растет признание того, что морские микроорганизмы играют гораздо более важную роль в морских экосистемах, чем считалось ранее. Развитие метагеномики дает исследователям возможность выявить ранее скрытые многообразия микроскопической жизни, предлагая мощную линзу для наблюдения за микробным миром и возможность революционизировать понимание живого мира. Методы диетического анализа с метабаркодированием используются для реконструкции пищевых цепей на более высоких уровнях таксономического разрешения и выявляют более глубокие сложности в паутине взаимодействий.
Микроорганизмы играют ключевую роль в морских пищевых сетях. Путь вирусного шунтирования — это механизм, который предотвращает миграцию микробных микробных частиц органического вещества (ПОМ) на трофические уровни за счет их рециркуляции в растворенное органическое вещество (РОВ), которое может легко поглощаться микроорганизмами. Вирусное шунтирование помогает поддерживать разнообразие в микробной экосистеме, предотвращая доминирование одного вида морских микробов в микросреде. РОВ, рециркулируемое путем вирусного шунтирования, сопоставимо с количеством, генерируемым другими основными источниками морского РОВ.
Как правило, растворенный органический углерод (DOC) попадает в окружающую среду океана в результате бактериального лизиса, утечки или экссудации фиксированного углерода из фитопланктона (например, слизистого экзополимера диатомовых водорослей ), внезапного клеточного старения, неаккуратного питания зоопланктоном, выделения отходов продукты, полученные водными животными, или разложение или растворение органических частиц наземных растений и почв. Бактерии в микробной петле разлагают этот детрит в виде частиц, чтобы использовать это богатое энергией вещество для роста. Поскольку более 95% органического вещества в морских экосистемах состоит из полимерных высокомолекулярных (ВМВ) соединений (например, белков, полисахаридов, липидов), только небольшая часть общего растворенного органического вещества (РОВ) может быть легко использована в большинстве морских экосистем. организмов на более высоких трофических уровнях. Это означает, что растворенный органический углерод недоступен для большинства морских организмов; морские бактерии вносят этот органический углерод в пищевую сеть, в результате чего дополнительная энергия становится доступной для более высоких трофических уровней.

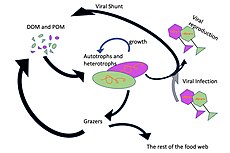



Гигантский вирус морской CroV заражает и вызывает смерть лизиса в морской zooflagellate кафетерий roenbergensis . Это влияет на экологию побережья, поскольку Cafeteria roenbergensis питается бактериями, обнаруженными в воде. Когда количество Cafeteria roenbergensis невелико из-за обширных инфекций CroV, бактериальные популяции растут в геометрической прогрессии. Влияние CroV на естественные популяции C. roenbergensis остается неизвестным; однако было обнаружено, что вирус очень специфичен для хозяина и не заражает другие близкородственные организмы. Cafeteria roenbergensis также инфицировано вторым вирусом, вирофагом Mavirus , который является сателлитным вирусом , что означает, что он может реплицироваться только в присутствии другого специфического вируса, в данном случае в присутствии CroV. Этот вирус препятствует репликации CroV, что приводит к выживанию клеток C. roenbergensis . Мавирус способен интегрироваться в геном клеток C. roenbergensis и тем самым обеспечивать иммунитет популяции.
Грибы


По среде обитания
Пелагические сети
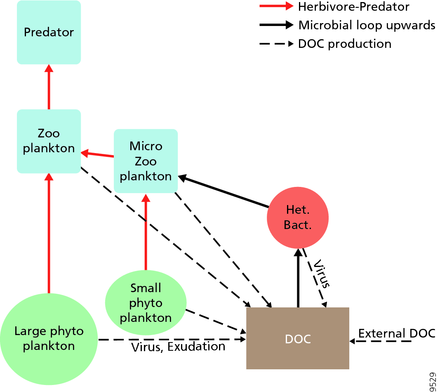
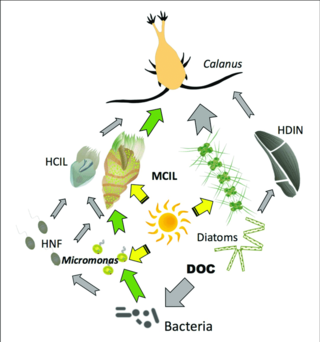
Для пелагических экосистем Лежандр и Расулзадаган предложили в 1995 году континуум трофических путей с пищевой цепью травоядных и микробной петлей в качестве конечных членов пищевой сети. Классический линейный конечный член пищевой цепи включает выпас зоопланктона на более крупном фитопланктоне и последующее нападение на зоопланктон либо более крупного зоопланктона, либо другого хищника. В такой линейной пищевой цепи хищник может либо привести к высокой биомассе фитопланктона (в системе с фитопланктоном, травоядным животным и хищником), либо к снижению биомассы фитопланктона (в системе с четырьмя уровнями). Таким образом, изменения численности хищников могут приводить к трофическим каскадам . Конечный член микробной петли включает не только фитопланктон в качестве основного ресурса, но и растворенный органический углерод . Растворенный органический углерод используется гетеротрофными бактериями для роста, которому предшествует более крупный зоопланктон. Следовательно, растворенный органический углерод через петлю бактерий-микрозоопланктон трансформируется в зоопланктон. Эти два конечных элемента обработки углерода связаны на нескольких уровнях. Мелкий фитопланктон может напрямую потребляться микрозоопланктоном.
Как показано на диаграмме справа, растворенный органический углерод производится разными способами и различными организмами, как первичными производителями, так и потребителями органического углерода. Высвобождение DOC первичными продуцентами происходит пассивно путем утечки и активно во время несбалансированного роста при ограничении питательных веществ. Другой прямой путь от фитопланктона к растворенному органическому бассейну — это вирусный лизис . Морские вирусы являются основной причиной смертности фитопланктона в океане, особенно в более теплых водах низких широт. Другими источниками растворенного органического углерода являются небрежное кормление травоядными животными и неполное переваривание добычи потребителями. Гетеротрофные микробы используют внеклеточные ферменты для растворения твердых частиц органического углерода и используют этот и другие ресурсы растворенного органического углерода для роста и поддержания. Часть микробной гетеротрофной продукции используется микрозоопланктоном; другая часть гетеротрофного сообщества подвергается интенсивному вирусному лизису, что снова вызывает высвобождение растворенного органического углерода. Эффективность микробной петли зависит от множества факторов, но, в частности, от относительной важности хищничества и вирусного лизиса для смертности гетеротрофных микробов.

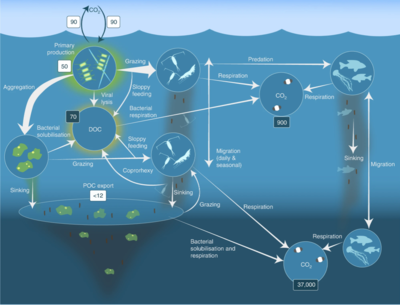
Пелагическая пищевая сеть и биологический насос. Связь между биологическим насосом океана и пелагической пищевой цепью, а также возможность удаленного отбора проб этих компонентов с кораблей, спутников и автономных транспортных средств. Светло-голубые воды — это эвфотическая зона , а более темно-синие воды — сумеречная зона .

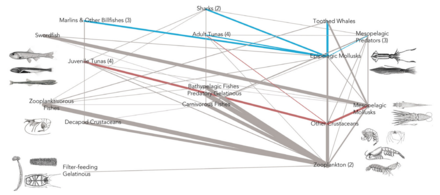
Влияние мезопелагических видов на глобальный углеродный бюджет
DVM = diel вертикальная миграция NM = немиграция

Мезопелагические щетиноротики могут быть самыми многочисленными позвоночными на планете, хотя о них мало что известно.

Студенистые хищники, подобные этому наркомедузану, потребляют наибольшее разнообразие мезопелагической добычи.
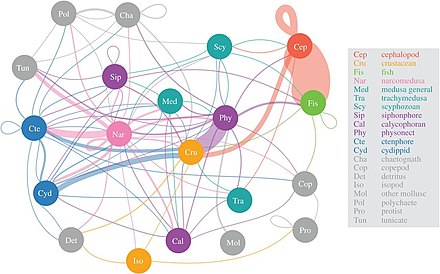
Ученые начинают более детально исследовать малоизвестную сумеречную зону мезопелагиали , глубиной от 200 до 1000 метров. Этот слой отвечает за удаление из атмосферы около 4 миллиардов тонн углекислого газа ежегодно. Мезопелагический слой населен большей частью биомассы морских рыб .
Согласно исследованию 2017 года, наркомедузы потребляют наибольшее разнообразие мезопелагических жертв, за ними следуют сифонофоры , гребневики и головоногие моллюски . Важность так называемой «желейной сети» только начинает осознаваться, но кажется, что медузы, гребневики и сифонофоры могут быть ключевыми хищниками в глубоких пелагических пищевых сетях с экологическими последствиями, подобными хищным рыбам и кальмарам. Традиционно студенистые хищники считались неэффективными поставщиками морских трофических путей, но они, по-видимому, играют существенную и неотъемлемую роль в глубоких пелагических пищевых сетях. Вертикальная миграция Диль , важный активный транспортный механизм, позволяет мезозоопланктону связывать углекислый газ из атмосферы, а также обеспечивать потребности в углероде других мезопелагических организмов.
В исследовании 2020 года сообщается, что к 2050 году глобальное потепление может распространяться в глубинах океана в семь раз быстрее, чем сейчас, даже если выбросы парниковых газов сократятся. Потепление в мезопелагических и более глубоких слоях может иметь серьезные последствия для глубоководной океанической пищевой сети, поскольку океанические виды должны будут перемещаться, чтобы оставаться при температурах выживания.
На поверхности океана

Среда обитания на поверхности океана находится на границе между океаном и атмосферой. В подобной биопленке среде обитания на поверхности океана обитают микроорганизмы, обитающие на поверхности, обычно называемые нейстоном . Эта обширная граница раздела воздух-вода находится на пересечении основных процессов обмена воздух-вода, охватывающих более 70% глобальной площади поверхности. Бактерии в поверхностном микрослое океана, называемые бактерионейстоном , представляют интерес из-за практических применений, таких как обмен парниковых газов между воздухом и морем, производство климатически активных морских аэрозолей и дистанционное зондирование океана. Особый интерес представляет производство и разложение поверхностно-активных веществ (поверхностно-активных материалов) посредством микробных биохимических процессов. Основные источники поверхностно-активных веществ в открытом океане включают фитопланктон, поверхностный сток и осаждения из атмосферы.
В отличие от цветения цветных водорослей, бактерии, связанные с поверхностно-активными веществами, могут быть не видны на цветных изображениях океана. Возможность обнаруживать эти «невидимые» бактерии, связанные с поверхностно-активными веществами, с помощью радара с синтетической апертурой дает огромные преимущества в любых погодных условиях, независимо от облачности, тумана или дневного света. Это особенно важно при очень сильном ветре, поскольку именно в этих условиях имеют место наиболее интенсивные газообмены между воздухом и морем и образование морских аэрозолей. Таким образом, в дополнение к цветным спутниковым изображениям, спутниковые изображения SAR могут дать дополнительную информацию о глобальной картине биофизических процессов на границе между океаном и атмосферой, обмене парниковыми газами между воздухом и морем и производстве климатически активных морских аэрозолей.
На дне океана
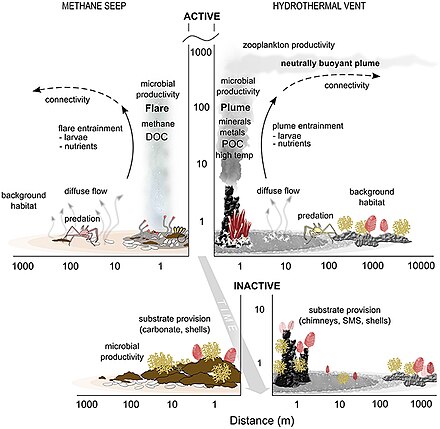
Среды обитания на дне океана ( бентосные ) находятся на границе между океаном и недрами земли.
Отводы и вентиляционные отверстия
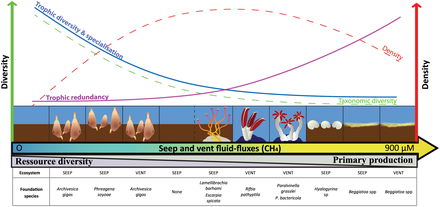
Прибрежные сети
Прибрежные воды включают воды в устьях и над континентальными шельфами . Они занимают около 8 процентов общей площади океана и составляют около половины всей продуктивности океана. Основными питательными веществами, определяющими эвтрофикацию, являются азот в прибрежных водах и фосфор в озерах. Оба они содержатся в высоких концентрациях в гуано (фекалиях морских птиц), которые служат удобрением для окружающего океана или соседнего озера. Мочевая кислота является доминирующим азотистым соединением, и во время ее минерализации образуются различные формы азота.
Экосистемы, даже с кажущимися четкими границами, редко функционируют независимо от других смежных систем. Экологи все больше осознают важное влияние межэкосистемного переноса энергии и питательных веществ на популяции и сообщества растений и животных. Хорошо известным примером этого является то, как морские птицы концентрируют питательные вещества морского происхождения на островах размножения в виде фекалий (гуано), которые содержат
15–20% азота (N), а также 10% фосфора. Эти питательные вещества резко изменяют функционирование и динамику наземных экосистем и могут способствовать повышению первичной и вторичной продуктивности. Однако, хотя многие исследования продемонстрировали обогащение наземных компонентов азотом из-за осаждения гуано в различных таксономических группах, лишь немногие изучали его влияние на морские экосистемы, и большинство этих исследований ограничивалось регионами с умеренным климатом и водами с высоким содержанием биогенных веществ. коралловые рифы могут быть найдены рядом с островами с большими популяциями гнездящихся морских птиц, и на них может потенциально повлиять местное обогащение питательными веществами из-за переноса питательных веществ, полученных из морских птиц, в окружающие воды. Исследования влияния гуано на тропические морские экосистемы показывают, что азот гуано обогащает морскую воду и первичных продуцентов рифов.
Кораллы, строящие рифы, имеют существенные потребности в азоте, и, процветая в бедных питательными веществами тропических водах, где азот является основным ограничивающим питательным веществом для первичной продуктивности, они разработали специальные приспособления для сохранения этого элемента. Их создание и поддержание частично связано с их симбиозом с одноклеточными динофлагеллятами, Symbiodinium spp. (зооксантеллы), которые могут поглощать и удерживать растворенный неорганический азот (аммоний и нитрат) из окружающих вод. Эти зооксантеллы также могут перерабатывать отходы животных и впоследствии передавать их обратно кораллам-хозяевам в виде аминокислот, аммония или мочевины. Кораллы также способны поглощать богатые азотом частицы отложений и планктон. Эвтрофикация прибрежных районов и избыток питательных веществ могут оказывать сильное воздействие на кораллы, приводя к замедлению роста скелета,
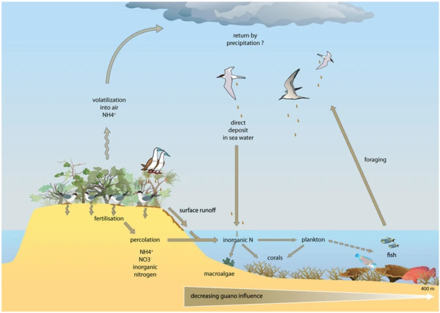
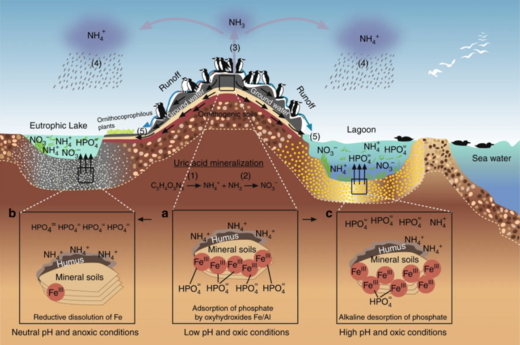
На диаграмме вверху справа: (1) при аммонификации образуются NH3 и NH4 +, и (2) при нитрификации образуется NO3- в результате окисления NH4 +. (3) в щелочных условиях, типичных для фекалий морских птиц, NH3 быстро улетучивается и превращается в NH4 +, (4) который выводится из колонии, а через влажные отложения экспортируется в отдаленные экосистемы, которые подвергаются эвтрофированию. Фосфорный цикл проще и имеет ограниченную подвижность. Этот элемент содержится в ряде химических форм в фекалиях морских птиц, но наиболее подвижным и биодоступным является ортофосфат (5), который может вымываться из подземных или поверхностных вод.

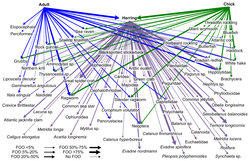
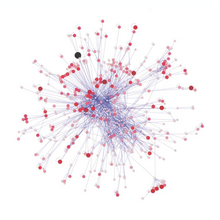
Штрих-кодирование ДНК можно использовать для построения структур пищевой сети с лучшим таксономическим разрешением в веб-узлах. Это обеспечивает более конкретную идентификацию видов и большую ясность в том, кто кого ест. «Штрих-коды ДНК и информация ДНК могут позволить новые подходы к построению более крупных сетей взаимодействия и преодолеть некоторые препятствия на пути к достижению адекватного размера выборки».
Недавно применяемый метод идентификации видов — это метабаркодирование ДНК . Идентификация видов по морфологии относительно сложна и требует много времени и опыта. Высокопроизводительное секвенирование. Метабаркодирование ДНК обеспечивает таксономическое назначение и, следовательно, идентификацию всего образца в отношении группоспецифичных праймеров, выбранных для предыдущей амплификации ДНК .

- Штрих-кодирование микробной ДНК
- Штрих-кодирование ДНК водорослей
- Штрих-кодирование ДНК рыб
- Штрих-кодирование ДНК при оценке диеты
- Ламинарии леса
- Бирнс, Дж. Э., Рейнольдс, П. Л. и Стахович, Дж. Дж. (2007) «Вторжения и вымирания меняют прибрежные морские пищевые сети». PLOS ONE , 2 (3): e295. DOI : 10.1371 / journal.pone.0000295
Полярные сети
Морские системы Арктики и Антарктики имеют очень разные топографические структуры и, как следствие, очень разные структуры пищевых сетей . Как арктические, так и антарктические пелагические пищевые сети имеют характерные потоки энергии, контролируемые в основном несколькими ключевыми видами. Но и для них не существует единой универсальной сети. Альтернативные пути важны для устойчивости и поддержания потоков энергии. Однако эти более сложные альтернативы обеспечивают меньший поток энергии к видам верхнего трофического уровня. «Структура пищевой сети может быть схожей в разных регионах, но отдельные виды, которые доминируют на средних трофических уровнях, различаются в полярных регионах».



Пищевая сеть Арктики сложна. Исчезновение морского льда может в конечном итоге затронуть всю пищевую сеть, от водорослей и планктона до рыб и млекопитающих. Воздействие изменения климата на конкретных видах может рябь через трофический и влияет на широкий спектр других организмов . Это не только снижение морского льда ухудшей популяции белого медведя путем уменьшения степени их первичного обитания, также негативно воздействуя на них через эффекты пищевой сети. Уменьшение продолжительности и протяженности морского льда в Арктике приводит к сокращению численности ледяных водорослей, которые процветают в богатых питательными веществами карманах во льду. Эти водоросли поедаются зоопланктоном, который, в свою очередь, поедает арктическая треска, важный источник пищи для многих морских млекопитающих, включая тюленей. Тюленей едят белые медведи. Следовательно, сокращение ледяных водорослей может способствовать сокращению популяций белых медведей.
В 2020 году исследователи сообщили, что измерения первичной продукции в Северном Ледовитом океане за последние два десятилетия показывают увеличение почти на 60% из-за более высоких концентраций фитопланктона . Они предполагают, что новые питательные вещества поступают из других океанов, и предполагают, что это означает, что Северный Ледовитый океан может поддерживать продукцию более высокого трофического уровня и дополнительную фиксацию углерода в будущем.




Антарктическая медуза Diplulmaris antarctica подо льдом

Колонии водоросли Phaeocystis antarctica , важного фитопланктера моря Росса, которая доминирует в начале сезона, цветет после отступления морского льда и выделяет значительное количество углерода.




Виды фундамента и краеугольного камня



Концепция основных видов была введена в 1972 году Полом К. Дейтоном , который применил ее к некоторым членам сообществ морских беспозвоночных и водорослей . Из исследований, проведенных в нескольких местах, стало ясно, что существует небольшая горстка видов, деятельность которых оказывает непропорционально большое влияние на остальную часть морского сообщества, и поэтому они являются ключом к устойчивости сообщества. Мнение Дейтона заключалось в том, что сосредоточение внимания на основных видах позволит использовать упрощенный подход для более быстрого понимания того, как сообщество в целом отреагирует на нарушения, такие как загрязнение, вместо того, чтобы пытаться выполнять чрезвычайно сложную задачу одновременного отслеживания реакции всех членов сообщества.
Основные виды — это виды, которые играют доминирующую роль в структурировании экологического сообщества , формировании его окружающей среды и определении его экосистемы. Такие экосистемы часто называют в честь основных видов, таких как луга морских водорослей, устричные русла, коралловые рифы, леса водорослей и мангровые леса. Например, красные мангровые заросли являются обычным основанием мангровых лесов. Корень мангрового дерева является местом разведения молодой рыбы, например, луциана . Основной вид может занимать любой трофический уровень в пищевой сети, но, как правило, является производителем.


Концепция краеугольного камня была введена в 1969 году зоологом Робертом Т. Пейном . Пейн разработал концепцию, чтобы объяснить свои наблюдения и эксперименты по взаимоотношениям между морскими беспозвоночными в приливной зоне (между линиями прилива и отлива), включая морских звезд и мидий . Некоторые морские звезды охотятся на морских ежей , мидий и других моллюсков , у которых нет других естественных хищников. Если морскую звезду удалить из экосистемы, популяция мидий бесконтрольно взорвется, вытеснив большинство других видов.
Ключевые виды — это виды, которые оказывают большое влияние, непропорционально своей численности, в пищевых сетях экосистемы. В экосистеме может произойти резкий сдвиг, если ключевой вид будет удален, даже если этот вид был небольшой частью экосистемы по показателям биомассы или продуктивности . Морские выдры ограничивают ущерб, который морские ежи наносят лесам ламинарии . Когда на каланов западного побережья Северной Америки охотились из-за их меха, их численность упала до такого низкого уровня, что они не смогли контролировать популяцию морских ежей. В ежи в свою очередь , паслись holdfasts из ламинарии так сильно , что бурых водорослей в значительной степени исчезли, наряду со всеми видами , которые зависят от них. Повторное введение каланов позволило восстановить экосистему водорослей.
Загадочные взаимодействия

Загадочные взаимодействия, взаимодействия, которые «скрыты на виду», происходят во всей морской планктонной пищевой сети, но в настоящее время в значительной степени игнорируются установленными методами, что означает, что сбор крупномасштабных данных для этих взаимодействий ограничен. Несмотря на это, текущие данные свидетельствуют о том, что некоторые из этих взаимодействий могут оказывать ощутимое влияние на динамику пищевой сети и результаты моделирования. Включение загадочных взаимодействий в модели особенно важно для взаимодействий, связанных с переносом питательных веществ или энергии.
Упрощения, такие как «зоопланктон потребляет фитопланктон», «фитопланктон поглощает неорганические питательные вещества», «валовая первичная продукция определяет количество углерода, доступного для пищевой сети» и т. Д., Помогли ученым объяснить и смоделировать общие взаимодействия в водной среде. Традиционные методы сосредоточены на количественной оценке и оценке этих обобщений, но быстрый прогресс в области геномики, пределов обнаружения сенсоров, экспериментальных методов и других технологий в последние годы показал, что обобщение взаимодействий внутри планктонного сообщества может быть слишком простым. Эти усовершенствования в технологии выявили ряд взаимодействий, которые кажутся загадочными, поскольку усилия по отбору массовых проб и экспериментальные методы направлены против них.
Сложность и стабильность
Пищевые сети обеспечивают основу, в рамках которой может быть организована сложная сеть взаимодействий хищник-жертва. Модель пищевой сети — это сеть пищевых цепочек . Каждая пищевая цепочка начинается с первичного продуцента или автотрофа , организма, такого как водоросль или растение, которое способно производить свою собственную пищу. Следующим в цепочке идет организм, который питается первичным продуцентом, и эта цепочка продолжается в виде цепочки последовательных хищников. Организмы в каждой цепи сгруппированы по трофическим уровням в зависимости от того, сколько звеньев они удалены от первичных продуцентов. Длина цепи, или трофический уровень, является мерой количества видов, встречающихся по мере того, как энергия или питательные вещества перемещаются от растений к высшим хищникам. Пищевая энергия перетекает от одного организма к другому, к следующему и так далее, при этом часть энергии теряется на каждом уровне. На данном трофическом уровне может существовать один вид или группа видов с одними и теми же хищниками и жертвами.
В 1927 году Чарльз Элтон опубликовал влиятельное обобщение об использовании пищевых сетей, в результате чего они стали центральной концепцией в экологии. В 1966 году интерес к пищевым сетям возрос после экспериментального и описательного исследования Роберта Пейна приливных берегов, предполагающего, что сложность пищевой сети является ключом к поддержанию видового разнообразия и экологической стабильности. Это и другие открытия побудили многих теоретиков-экологов, в том числе Роберта Мэя и Стюарта Пимма , изучить математические свойства пищевых сетей. Согласно их анализу, сложные пищевые сети должны быть менее стабильными, чем простые пищевые сети. Очевидный парадокс между сложностью пищевых сетей, наблюдаемых в природе, и математической хрупкостью моделей пищевых сетей в настоящее время является областью интенсивных исследований и дискуссий. Парадокс может быть частично обусловлен концептуальными различиями между устойчивостью пищевой сети и равновесной стабильностью пищевой сети.
Трофический каскад может иметь место в пищевой сети , если трофический уровень в сети подавляется.
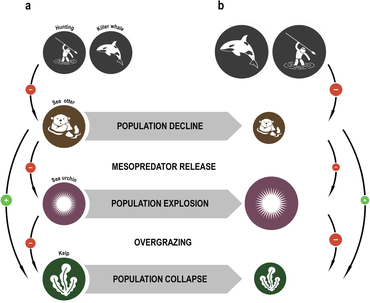
Например, нисходящий каскад может возникнуть, если хищники достаточно эффективны в хищничестве, чтобы уменьшить численность или изменить поведение своей жертвы , тем самым освобождая следующий более низкий трофический уровень от хищников. Нисходящий каскад — это трофический каскад, в котором главный потребитель / хищник контролирует популяцию основных потребителей . В свою очередь, население первичных производителей процветает. Удаление главного хищника может изменить динамику трофической сети. В этом случае первичные потребители будут перенаселенными и будут эксплуатировать первичных производителей. В конце концов, не хватит первичных производителей, чтобы поддерживать потребительское население. Стабильность нисходящей пищевой сети зависит от конкуренции и хищничества на более высоких трофических уровнях. Инвазивные виды также могут изменить этот каскад, удалив или превратившись в главного хищника. Это взаимодействие не всегда может быть отрицательным. Исследования показали, что некоторые инвазивные виды начали менять каскады; и, как следствие, была восстановлена деградация экосистемы. Пример каскада в сложной экосистеме открытого океана произошел в северо-западной части Атлантического океана в 1980-х и 1990-х годах. Изъятие атлантической трески ( Gadus morhua ) и других наземных рыб в результате длительного перелова привело к увеличению численности кормовых видов этих наземных рыб, особенно мелких кормовых рыб и беспозвоночных, таких как северный снежный краб ( Chionoecetes opilio ) и северные креветки. ( Pandalus borealis ). Увеличение численности этих видов добычи изменило сообщество зоопланктона, который служит пищей для более мелких рыб и беспозвоночных в качестве косвенного эффекта. Нисходящие каскады могут быть важны для понимания побочного эффекта удаления высших хищников из пищевых сетей, как это сделали люди во многих местах посредством охоты и рыбалки .
В восходящем каскаде популяция первичных продуцентов всегда будет контролировать увеличение / уменьшение энергии на более высоких трофических уровнях. Основными продуцентами являются растения, фитопланктон и зоопланктон, которым необходим фотосинтез. Хотя свет важен, популяция основных продуцентов изменяется в зависимости от количества питательных веществ в системе. Эта пищевая сеть зависит от доступности и ограниченности ресурсов. Все популяции будут расти, если изначально имеется большое количество питательных веществ.
Источник